
Эволюционное место и происхождение вторичноротых животных
Филогенетическое дерево вторичноротых животных
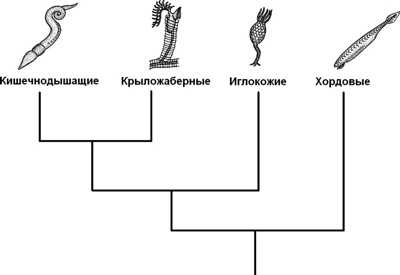 |
Раньше считалось, что в группу вторичноротых входят 3 типа животных:
Теперь в объединение вторичноротых включают также щетинкочелюстных и погонофор, |
Вторичноротые животные (дейтеростомы)
/ | |
/ / \ / \
/ / \ / \
Щетинкочелюстные Полухордовые Иглокожие Погонофоры Хордовые
|
|-перистожаберные (крыложаберные)
|
|-кишечнодышащие
|
|-граптолиты (ископаемые)
|
Гипотезы происхождения вторичноротых животных
|
Происхождение вторичноротых неясно.
У вторичноротых в период зародышевого развития на месте первичного рта (бластопора) образуется анальное отверстие,
Все билатерально-симметричные организмы делятся на две группы — первичноротые и вторичноротые. Так, у первичноротых животных бластопор (возникающее в эмбриональном развитии отверстие первичного кишечника) У вторичноротых (например, у иглокожих) бластопор становится анальным отверстием, Помимо судьбы рта, первичноротые и вторичноротые отличаются другими эмбриологическими особенностями. (1) Во-первых, это характер дробления оплодотворенного яйца: (2) Другое отличие состоит в способе закладке целома (вторичной полости тела): (3) Третьим важнейшим различием между первично- и вторичноротыми является судьба зачатка первичного мозга: |

Высказана гипотеза о происхождении вторичноротых от первичноротых.
Образ жизни и некоторые черты строения предка вторичноротых
Общий предок вторичноротых животных, по-видимому, вел донный образ жизни,
ползая на брюшной стороне с помощью ресничной полоски, подостланной вентральным нервным тяжем.
Он имел три пары целомических мешков и щупальцевый аппарат, с помощью которого собирал мелкие пищевые частицы.
В дальнейшем могло произойти обособление самого заднего отдела тела,
с помощью которого предки вторичноротых временно зарывались в верхние слои грунта
В эволюции вторичноротых этому отделу принадлежит очень важное место.
Очевидно, у него уже были особые отверстия, соединяющие полость глотки с внешней средой: жаберные поры.
Он был фильтратором и питался бактериями и органической взвесью, которая попадала в глотку с водой через рот
и осаждалась на слизистом эпителии, а вода выходила наружу через жаберные поры.
Именно так и сегодня живут примитивные вторичноротые (кишечнодышащие, ланцетник, личинки миног),
и такой же способ питания предполагается для древнейших ископаемых вторичноротых, таких как юннанозои и пикайя.
Особенности строения
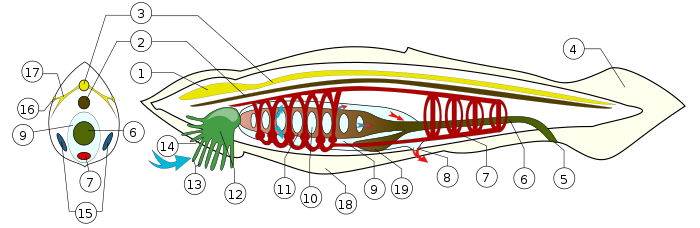 Схема строения головохордовых (b) на примере ланцетника (b) : 1 — утолщение нервной трубки (b) спереди («мозг (b) »); 2 — хорда (b) ; 3 — спинной нервный тяж («спинной мозг (b) »); 4 — хвостовой плавник; 5 — анус (b) ; 6 — пищеварительный канал (b) ; 7 — кровеносная система (b) ; 8 — выходное отверстие околожаберной полости (атриопор); 9 — околожаберная полость; 10 — глоточные (жаберные (b) ) щели; 11 — глотка (b) ; 12 — ротовая полость (b) ; 13 — околоротовые щупальца (b) ; 14 — ротовое отверстие (b) ; 15 — половые железы (b) (семенники (b) или яичники (b) ); 16 — глазки Гессе (b) ; 17 — нервы (b) ; 18 — метаплевральная складка; 19 — слепой печёночный вырост (b) . Дыхание (b) (газообмен (b) ): синей стрелкой указан вход воды, богатой кислородом (b) , а красными — выход обогащённой углекислым газом (b) .
Схема строения головохордовых (b) на примере ланцетника (b) : 1 — утолщение нервной трубки (b) спереди («мозг (b) »); 2 — хорда (b) ; 3 — спинной нервный тяж («спинной мозг (b) »); 4 — хвостовой плавник; 5 — анус (b) ; 6 — пищеварительный канал (b) ; 7 — кровеносная система (b) ; 8 — выходное отверстие околожаберной полости (атриопор); 9 — околожаберная полость; 10 — глоточные (жаберные (b) ) щели; 11 — глотка (b) ; 12 — ротовая полость (b) ; 13 — околоротовые щупальца (b) ; 14 — ротовое отверстие (b) ; 15 — половые железы (b) (семенники (b) или яичники (b) ); 16 — глазки Гессе (b) ; 17 — нервы (b) ; 18 — метаплевральная складка; 19 — слепой печёночный вырост (b) . Дыхание (b) (газообмен (b) ): синей стрелкой указан вход воды, богатой кислородом (b) , а красными — выход обогащённой углекислым газом (b) .
Хордовые — тип животных, характеризующихся билатеральной симметрией (b) и наличием, по крайней мере, на определённых стадиях развития следующих признаков:
Эволюция хордовых животных
Все началось с появления ланцетников, они — переходная форма между кольчатыми червями и позвоночными животными. У ланцетника, как и кольчатых червей, нет мозга и сердца. Мышцы червя поделены на сегменты специальными лентами. У ланцетника так же. А еще выделительные системы идентичны. О хордовых особенностях ланцетника говорилось выше в статье.
Наличие признаков двух разных групп животных — свидетельство переходной формы, как и данные ДНК-тестов. От кольчатых червей ланцетники отщепились около 500 миллионов лет назад. Тогда же стали отделяться позвоночные. Их предки выбрали хищнически образ жизни. Это обязывало к подвижности. Поэтому спинная струна преобразовалась в хрящ, а потом и в кость. Так скелет хордовых животных стал совсем не хордовым.
Следующий этап — формирование парных плавников. Это дало дополнительную скорость. Хищники с плавниками имели преимущество. Так появились рыбы и именно поэтому стали столь многочисленны. Быстрые движения развили нервную систему и органы чувств. Появились острые зубы. Моря наполнились монстрами до такой степени, что некоторым захотелось сбежать, даже самим монстрам.
Так плавники стали преобразовываться в лапы. Вышедшие на сушу рыбы стали земноводными. У них появилось 2 круга обращения крови и легочная система дыхания. Уходя все дальше от водоемов, земноводные потеряли кожное дыхание и стали пресмыкающимися. Вдали от воды, они стали и размножаться на суше, откладывать яйца. Так выглядит эволюция хордовых в соответствии с теорией Дарвина.
Млекопитающим пресмыкающиеся уступили, поскольку выработанное новыми животными теплокровие дало преимущество. Можно было расселиться в разные климатические зоны и адаптироваться к изменениям температурного режима на планете. Еще веским преимуществом млекопитающих стала выработанная ими забота о потомстве. Поэтому число рептилий стало сокращаться, а высшие позвоночные, наоборот, активно развивались, расселялись.
Сравнение хордовых с другими вторичноротыми
Хотя
хордовые — это вторичноротые животные, все же их развитие имеет ряд
особенностей. На рис. 3 представлены соответствующие стадии развития одной из
групп вторичноротых — погонофор и низших хордовых — асцидий.
Анимальный
полюс яйца погонофор, на котором располагается ядро яйцеклетки, соответствует
спинной стороне эмбриона, а вегетативный, на котором концентрируется желток, —
брюшной. У низших хордовый ситуация обратная: анимальный полюс соответствует
брюшной стороне, а вегетативный — спинной. Бластопор (то есть та область
эмбриона, где впячивается энтодерма и формируется первичный кишечник) у
погонофор соответствует брюшной стороне личинки, а у низших хордовых — спинной.
Зачаток нервной системы, закладывающийся по линии замыкания бластопора, у
погонофор становится брюшным нервным тяжем, а у хордовых — спинной нервной
трубкой (рис. 3).
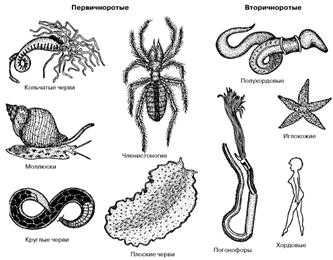
Рис.
2. Представители основных групп первичноротых и вторичноротых животных.
План
строения низших хордовых также существенно отличается от планов строения других
вторичноротых. Так, например, у всех вторичноротых передние целомы
диссимметричны: левый передний целом больше правого, который обычно выполняет
функцию околосердечной сумки. У хордовых ситуация обратная: правый передний
целом больше левого. Целомы вторичноротых сообщаются с внешней средой
отверстиями, открывающимися на спинной стороне и выполняющими осморегуляторные
и половые функции. У хордовых эти отверстия располагаются на той стороне тела,
которую мы традиционно считаем брюшной.
Кровеносная
система у вторичноротых и хордовых устроена по общему плану и состоит из двух
главных сосудов: спинного и брюшного. У вторичноротых, как и всех других
беспозвоночных, кровь по спинному сосуду течет вперед, а по брюшному назад. У
хордовых направление тока крови обратное: по спинному сосуду кровь течет назад,
а по брюшному — вперед. Сердце у вторичноротых лежит на спинной стороне, а у
хордовых — на брюшной.
Суммируем
все эти отличия.
1.
Бластопор хордовых соответствует не брюшной, а спинной стороне.
2.
Нервная пластинка хордовых, закладывающаяся по месту замыкания бластопора,
превращается не в брюшной нервный тяж, а в спинную нервную трубку.
3.
Диссимметрия первой пары целомов имеет обратный знак в сравнении с другими
вторичноротыми: правый передний целом у хордовых больше левого.
4.
Отверстия целомов у хордовых открываются не на спинной, а на брюшной стороне.
5.
Кровь у хордовых по брюшной стороне течет вперед, а по спинной — назад, и
сердце располагается не на спинной, а на брюшной стороне.
Что
же означают эти несоответствия? Только одно: предки хордовых в своем
эволюционном развитии испытали инверсию сторон тела, то есть перевернулись и
стали передвигаться на морфологически спинной стороне, которая стала
функционировать как физиологически брюшная. Морфологически брюшная сторона
предков хордовых стала функционировать как физиологически спинная.
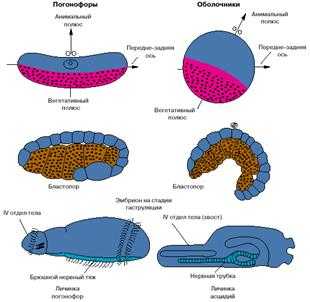
Рис.
3. Особенности эмбрионального развития погонофор и низших хордовых.
Если
мы примем этот вывод, все перечисленные выше несоответствия исчезнут: бластопор
хордовых станет соответствовать их исходно брюшной стороне, нервная трубка
будет располагаться на морфологически брюшной стороне, диссимметрия передних
целомов приобретет нормальный для вторичноротых характер, отверстия целомов
окажутся на морфологически спинной стороне, и направление тока крови, так же
как положение сердца, окажется типичным для вторичноротых. На рис. 4
представители низших хордовых (личинка асцидий и ланцетник) изображены
перевернутыми, чтобы продемонстрировать, что при таком положении их системы
органов соответствуют таковым других вторичноротых.
Переворот
на спинную сторону мог стать тем важнейшим эволюционным событием, которое
определило формирование плана строения хордовых животных и его коренное отличие
от планов строения вторичноротых и других беспозвоночных.
Происхождение
хордовых животных. В. В. МАЛАХОВ. Продолжение.
Особенности строения хордовых
Какие особенности строения сформировались у хордовых животных в процессе эволюции?
Кожные покровы у Бесчерепных состоят из однослойного эпителия, у позвоночных это пух, перья, чешуя, еще из производных эпидермиса – когти, рога, копыта. Появляются железы выделяющие пот, кожное сало, пахучие вещества. Например, у коров, коз, овец образуются молочные железы для кормления потомства.
Филогенез кровеносной системы хордовых идет из мезодермы, формируется круг кровообращения (замкнутый), сердце в брюшной области, спинная и брюшная артерии, которые соединены анастомозами. Стенки сосудов высланы эндотелиальными клетками, что способствует поддержанию гомеостаза, увеличилось количество глюкозы крови и белков (зачатки иммунитета).
Нервная система в эмбриональном периоде выглядит, как слой утолщенной эктодермы, окончания ее замыкаются и формируют нервную трубку. Филогенез головного мозга хордовых происходит из переднего конца нервной трубки, спинного – из заднего. Формирование головного мозга (кефализация) – стало необходимым условием эволюционного развития Хордовых. Они начали различать внешние раздражители, воспринимать окружающий мир с помощью органов чувств. Высшие хордовые имеют головной мозг с пятью отделами, которые сформировались из трех мозговых пузырей.
Развитие дыхательной системы и пищеварительной тесно связано, идет общая закладка под видом прямой трубки, которая у высших хордовых дифференцируется на две самостоятельные системы.
У низших хордовых пищеварительный тракт не имеет отделов, у ланцетников ротовое отверстие с множеством щупалец сообщается с глоткой, где расположен фильтрующий аппарат, пища затем через заднюю бороздку направляется в кишку и выводится анальным отверстием, железы не развиты. У позвоночных пищеварительная система уже состоит из отдельных структур: глотки, пищевода, желудочно-кишечного тракта, появляются секретирующие железы: печень и поджелудочная железа.
Дыхание осуществляется жабрами и легкими (наземные позвоночные). После выхода из воды у земноводных сформировались легкие, но функциональные возможности их недостаточны, поэтому дополнительный газообмен осуществляется через кожу. Таким образом, для Класса Земноводных характерно кожно-легочное дыхание.
Для класса Птиц характерно наличие двойного дыхания. Дыхательный аппарат состоит из гортани, трахеи, бронхов, легких с воздушными мешками. Во время полета птицы используют для газообмена воздушные мешки, вдыхаемый воздух дважды проходит через легкие – и на вдохе, и на выдохе – это и есть механизм двойного дыхания.
Система размножения у хордовых животных представлена семенниками и семяпроводами у самцов, яичниками и яйцепроводами у женских особей. Хордовые, за исключением личиночнохордовых, раздельнополые.
Примеры хордовых животных
Для простоты, приведем по паре примеров, разделив их, как и полагается систематикой, на подтипы.
Представители Бесчерепного подтипа
Европейский ланцетник
В длину животное до 8 сантиметров. Ткани белесые, слегка сероватые или желтоватые. Имеет промысловое значение за счет легкости употребления, ведь нет костей и питательности. В мясе много белка и полезных жирных кислот, в том числе, редкой доказопентоеновой кислоты. Среди ланцетников Европейский саамы известны и распространенный. Описан в 18 веке, как моллюск. Потом, животное причислили к рыбам. Правильно классифицировали только в 20 столетии.
Азиатский ланцетник
Этот вид промысловый. В Азии попадает в суп и жаркое. Еще ланцетников сушат для экспорта. Такие снеки заказывают в Сицилии, Сингапуре и на острове Ява. На помысел за ланцетником выходят на юго-западе Восточно-Китайского моря. В КНР азиатский вид именую рыбой бога литературы. Согласно мифам, тысячи хордовых вышли после смерти из дракона, на котором Бог литературы путешествовал на небо. В длину представители вида достигают 10 сантиметров и отличается красноватым цветом.
Представители Оболочников
Асцидия Ботриллул Бугорчатый
Относится к живущим колониями. Вся колония в длину не превышает 5 сантиметров. Соответственно, отдельные асцидии еще миниатюрнее. Есть подвиды с матовой и полностью прозрачной туникой. Она окрашена в оттенки фиолетового. Туника Ботриллула Бугорчатого активно пронизана сосудами. Они просвечивают.
Аппендикулярия Ойкоплевра Двудомная
Единственный раздельнополый вид. Отличается и расположение половых гамет в теле. У Ойкоплевры Двудомной они в задней части тела. Представители вида вытянуто-овальной формы и отличаются большим эндостилем, доходящим почти до анального отверстия.
Представители подтипа Позвоночные
Щелезуб
Относится к млекопитающим отряда насекомоядных. Щелезуб достигает полуметра в длину с учетом 25-сантиметрового хвоста. Весит зверек около 1 кило и отличается экзотической внешностью. Выделяется, к примеру, длинный нос-мордочка, помогающий хордовому млекопитающему животному добывать пропитание.
Трубкозуб
Напоминает свинью с удлиненной мордой и заячьими ушами. Относится к млекопитающим. Животное почти голокожее, обитает в Африке где именуется аардварк. На местном это «земляная свинья». Почему свинья, разобрались. Земляная же она, поскольку забирается в норы. Сзади у трубкозуба мускулистый хвост, словно взятый у кенгуру.
Исполинская саламандра
Самая большая среди саламандр планеты. Родина — страна Восходящего Солнца. Японская саламандра — земноводное длиною до 1,6 метра. Упитана. При такой длине может быть 180 кило. Внушителен и срок жизни. Предел — 1,5 века.
Суматранский носорог
Самый миниатюрный из носорогов. Живет на острове Суматра. Отсюда название вида. Взрослый суматранец не превышает в длину 2,8 метра. Чаще, это 1,5-2 метра. Высота животного достигает 150 сантиметров, но чаще, останавливается на 1 метре. Вес суматранского носорога в пределах 1 тонны.
Рыба Мола Мола
Иначе именуется Луной за схожесть в цвете и форме. Рыба уплощенная, словно камбала, но гораздо больше. В длину животное достигает 500 сантиметров. Весит Мола Мола до 1,5 тонн. Такую поймали в Америке. Идут споры об образе жизни рыбы Луны. Взрослых особей ловят у поверхности, где их отлично видно. В желудках добытых особей ничего не находят. Отсюда предположение, что всплывают только больные рыбы. Возможно, Мола Мола — глубоководный вид.
Фосса
Водится только на Мадагаскаре. Является млекопитающим и относится к кошачьим. Фосса — самый крупный хищник острова. Длина тела — 0,8 метра. Плюсом такой же хвост. У Фоссы приземистое, крепкое сложение, но длинные лапы.
Происхождение
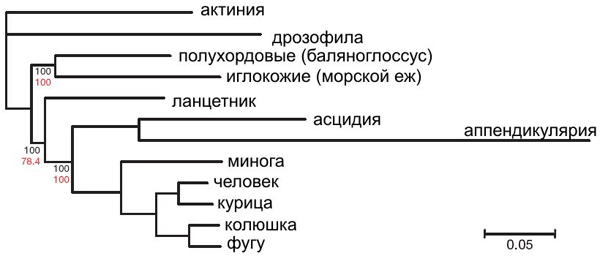
Попытки выработать эволюционные отношения хордовых привели к рождению нескольких гипотез. В настоящее время достигнут консенсус в том, что хордовые являются потомками одного общего предка, который сам является хордовым, а ближайшими родственниками позвоночных (лат. Vertebrata) являются головохордовые (лат. Cephalochordata). Все обнаруженные окаменелости ископаемых хордовых были найдены в раннем кембрии и включают в себя два вида позвоночных, классифицируемых как рыба. Поскольку окаменелости хордовых плохо сохранились, только метод молекулярной филогенетики предполагает разумную перспективу исследования их возникновения. Однако, использование метода молекулярной филогенетики для изучения эволюционных процессов является спорным.
Билатеральные животные делятся на два больших таксона — первичноротые и вторичноротые. Хордовые относятся ко вторичноротым. Весьма вероятно, что ископаемое кимберелла, жившая 555 млн лет назад, принадлежала к первичноротым. Жившая 549—543 млн лет назад в эдиакарии эрниетта была уже явно вторичноротым животным. Таким образом, первичноротые и вторичноротые должны были разделиться до времени существования этих животных, то есть до начала кембрийского периода.
Первые известные ископаемые двух близких к хордовым групп — иглокожих и полухордовых — обнаруживаются с раннего и среднего кембрия, соответственно. Кроме того известно ископаемое чэнцзянской биоты Yunnanozoon
, точная принадлежность которого к полухордовым или хордовым не определена. Другое ископаемое, относящееся к той же биоте,Haikouella lanceolata явно является хордовым и, возможно, позвоночным. У него обнаружены признаки сердца, артерий, нервной трубки и мозга, хвоста, жаберных лепестков, возможно, глаз, но в то же время вокруг ротового отверстия есть щупальца.Haikouichthys иMyllokunmingia , также из шенженьгской фауны и пикайя из сланцев Бёрджес относятся к примитивным позвоночным. С другой стороны, ископаемые других хордовых весьма редки, поскольку у них нет твёрдых частей тела.
Исследования родственных отношений хордовых начались с 90-х годов XIX века. Они основывались на анатомических, эмбриологических и палеонтологических данных и приводили к разным филогенетическим деревьям. Некоторое время ближайшими родственниками хордовых считали полухордовых, но ныне эта гипотеза отвергнута. Сочетание данных классических методов с данными по анализу последовательностей генов рРНК привело к появлению гипотезы о том, что оболочники — живые представители группы, базальной для других вторичноротых. Относительно взаимоотношений внутри хордовых некоторые учёные считают, что наиболее близкими родственниками позвоночных являются головохордовые, но есть основания считать таковыми оболочников.
Время происхождения хордовых, на основании метода молекулярных часов, было оценено в 896 млн лет.
Ниже приведена кладограмма, основанная на сравнении последовательностей митохондриальных ДНК:
| Deuterostomia | (Вторичноротые) | Xenoturbellida | |
| Hemichordata (Полухордовые) | |||
| Echinodermata (Иглокожие) |
Chordata (Хордовые)
| Cephalochordata (Бесчерепные) | |
| Tunicata (Urochordata, Оболочники) | |
| Craniata (Черепные) |
Систематика Хордовых
Тип Хордовые делится на три подтипа.
Подтип Оболочники (Личиночнохордовые) насчитывает примерно 1000 видов, имеющих хорду лишь в начальных стадиях развития.
Класс асцидий ведет сидячий образ жизни, они цепляются к камням на дне моря, или к основанию судов, образуя сплошные пласты из мелких особей. Асцидии имеют отверстия: ротовое служит фильтром для захваченной воды, отфильтрованные пища поступает в кишечник и затем продукты переваривания выводятся через анальное отверстие.
Сальпы и Аппендикулярии способны свободно перемещаться в водных просторах.
Оболочники гермафродиты (присутствуют и семенники, и яичники), из яиц выходят личинки, которые мало похожи на взрослую особь, имеют признаки внутреннего скелета, что свидетельствует о родстве с позвоночными.
Подтип Бесчерепные – морские обитатели придонной территории, сохранили в себе признаки типичных Хордовых (нервная трубка, жабры, хорда). Головной мозг не развит, ведут примитивный образ существования, достигая максимальной величины в 7см, представители раздельнополые особи, мечут икру, живут в песках на дне морских глубин и океанов, оставив снаружи только часть тела с ротовой полостью для питания. Выделяют один Класс Ланцетники.
Подтип Позвоночные – высокоорганизованные особи, с наличием развитых полушарий головного мозга и черепной коробки, позвоночного столба, который служит защитной оболочкой для спинного мозга. В процессе эволюции получили сложное морфологическое строение, усовершенствованные органы чувств, перешли от сидячего образа жизни, стали легко перемещаться по просторам земли, воздуха и воды. Позвоночные включают земноводных, рыб, птиц, пресмыкающихся, млекопитающих и круглоротых.
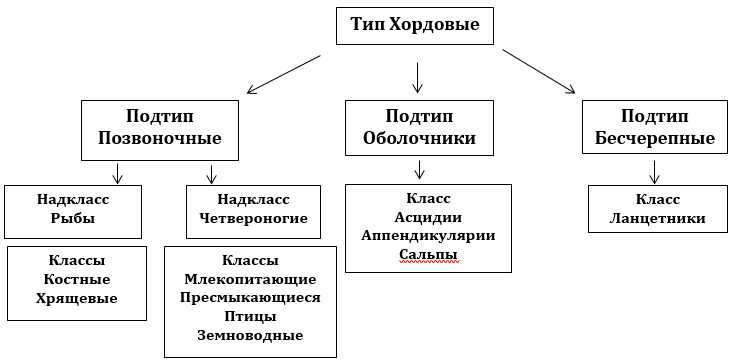 Классификация Хордовых — схема
Классификация Хордовых — схема
Тип Хордовые (chordata) — примеры животных
Представители хордовых животных:
-
рыбы любых видов, от речного карася до акулы;
- земноводные: жаба, лягушка, тритон;
I
-
пресмыкающиеся: ящерица, черепаха, змея, крокодил, хамелеон;
-
птицы: начиная с отряда аистообразных (аист, ибис цапля), буревестникообразных (альбатрос, буревестник), заканчивая воробьинообразными (жаворонок, ласточка, трясогузка, королек, дрозд, синица, скворец, воробей) и гусеобразными (гусь, утка, лебедь);
-
млекопитающие: семейство псовые (волк, лисица, собака и т. д.), семейство кошачьи (лев, тигр, гепард, рысь), семейство медвежьи (медведь, коала, панда).
https://youtube.com/watch?v=MzHptfLX7KY
Заключение
Современные
представления о происхождении хордовых объединяют положения, существовавшие
ранее в рамках альтернативных гипотез происхождения хордовых: перевернутость
хордовых, их родство с вторичноротыми и факт большей примитивности личинок
оболочников по сравнению с другими хордовыми. В отличие от ранее существовавших
точек зрения хордовые рассматриваются не как перевернутые кольчатые черви и
членистоногие, но как перевернутые вторичноротые. Что же касается личинок
асцидий, то они не являются предками остальных хордовых, как это принимается в
гипотезах неотенического происхождения хордовых. Организация личинок асцидий
рекапитулирует (то есть повторяет в сжатой, видоизмененной форме) в индивидуальном
развитии определенный филогенетический этап в историческом развитии хордовых, а
именно строение предков хордовых до их переворота.
Дискуссия
о происхождении хордовых не закончена. Развитие биологической науки приносит
новые доводы в пользу той или иной точки зрения. Иногда эти доводы приходят с
совсем неожиданной стороны. Так, относительно недавно, в 1993 году, в
английском журнале «Nature» появилась статья биологов Слэка, Холланда
и Грэма, анализирующая многочисленные публикации, в которых были выявлены зоны
экспрессии одной интересной группы генов (так называемой Hox-группы) в развитии
различных беспозвоночных и позвоночных животных. В этих публикациях было
показано, что у беспозвоночных эти гены маркируют брюшную сторону, а у
позвоночных их экспрессия наблюдается на спинной стороне. Этот факт хорошо
согласуется с идеей перевернутости хордовых, принимаемой современной
сравнительной анатомией.